Medikal na dalubhasa ng artikulo

Mga bagong publikasyon
Obaryo
Huling nasuri: 04.07.2025

Ang lahat ng nilalaman ng iLive ay medikal na nasuri o naka-check ang katotohanan upang masiguro ang mas tumpak na katumpakan hangga't maaari.
Mayroon kaming mahigpit na mga panuntunan sa pag-uukulan at nag-uugnay lamang sa mga kagalang-galang na mga site ng media, mga institusyong pang-akademikong pananaliksik at, hangga't maaari, ang mga pag-aaral ng medikal na pag-aaral. Tandaan na ang mga numero sa panaklong ([1], [2], atbp) ay maaaring i-click na mga link sa mga pag-aaral na ito.
Kung sa tingin mo na ang alinman sa aming nilalaman ay hindi tumpak, hindi napapanahon, o kung hindi pinag-uusapan, mangyaring piliin ito at pindutin ang Ctrl + Enter.
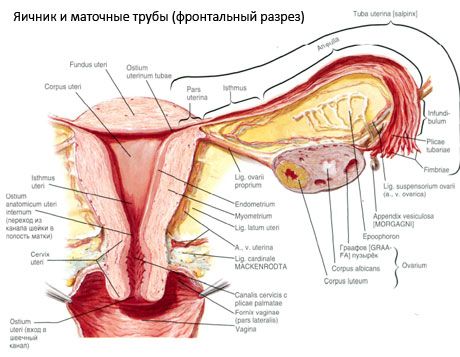
Ang ovary (ovarium; Greek oophoron) ay isang nakapares na organ, isang babaeng reproductive gland, na matatagpuan sa pelvic cavity sa likod ng malawak na ligament ng matris. Sa mga ovary, ang mga babaeng reproductive cell (mga itlog) ay bubuo at nagiging mature, at ang mga babaeng sex hormone ay nabuo na pumapasok sa dugo at lymph. Ang obaryo ay may hugis-itlog na hugis, medyo patag sa anteroposterior na direksyon. Ang kulay ng obaryo ay pinkish. Sa ibabaw ng obaryo ng isang babae na nanganak, makikita ang mga depresyon at mga peklat - mga bakas ng obulasyon at ang pagbabago ng corpora lutea. Ang obaryo ay tumitimbang ng 5-8 g. Ang mga sukat ng obaryo ay: haba 2.5-5.5 cm, lapad 1.5-3.0 cm, kapal - hanggang 2 cm. Ang obaryo ay may dalawang libreng ibabaw: ang medial surface (facies medialis), nakaharap sa pelvic cavity, bahagyang sakop ng fallopian tube, at ang lateral surface (facies lateralis), na katabi ng side wall ng pelvis, sa isang bahagyang ipinahayag na depression - ang ovarian fossa. Ang fossa na ito ay matatagpuan sa anggulo sa pagitan ng mga panlabas na iliac vessel na natatakpan ng peritoneum sa itaas at ang uterine at obturator arteries sa ibaba. Sa likod ng obaryo, ang ureter ng kaukulang panig ay dumadaan nang retroperitoneally mula sa itaas hanggang sa ibaba.
Ang mga ibabaw ng ovary ay pumasa sa isang convex free (posterior) na gilid (margo liber), sa harap - sa mesenteric edge (margo mesovaricus), na nakakabit sa pamamagitan ng isang maikling fold ng peritoneum (mesentery ng ovary) sa posterior leaflet ng malawak na ligament ng matris. Sa anterior na gilid ng organ ay mayroong isang grooved depression - ang hilum ng ovary (hilum ovarii), kung saan ang arterya at nerbiyos ay pumapasok sa obaryo, at ang mga ugat at lymphatic vessel ay lumabas. Ang obaryo ay mayroon ding dalawang dulo: ang bilugan na itaas na tubular na dulo (extremitas tubaria), nakaharap sa fallopian tube, at ang lower uterine end (extremitas utenna), na konektado sa matris ng ovarian ligament (lig. ovarii proprium). Ang ligament na ito, sa anyo ng isang bilog na kurdon na halos 6 mm ang kapal, ay napupunta mula sa dulo ng matris ng obaryo hanggang sa lateral na anggulo ng matris, na matatagpuan sa pagitan ng dalawang dahon ng malawak na ligament ng matris. Kasama rin sa ligamentous apparatus ng ovary ang ligament suspensory ng ovary (lig.suspensorium ovarii), na isang fold ng peritoneum na tumatakbo mula sa dingding ng maliit na pelvis hanggang sa ovary at naglalaman ng mga ovarian vessel at bundle ng fibrous fibers sa loob. Ang obaryo ay naayos ng isang maikling mesentery (mesovarium), na isang duplikasyon ng peritoneum na tumatakbo mula sa posterior leaflet ng malawak na ligament ng matris hanggang sa mesenteric na gilid ng obaryo. Ang mga ovary mismo ay hindi sakop ng peritoneum. Ang pinakamalaking ovarian fringe ng fallopian tube ay nakakabit sa tubular na dulo ng obaryo. Ang topograpiya ng obaryo ay nakasalalay sa posisyon ng matris, laki nito (sa panahon ng pagbubuntis). Ang mga ovary ay napaka-mobile na organo ng maliit na pelvic cavity.
 [
[ Mga daluyan at nerbiyos ng obaryo
Ang suplay ng dugo sa mga obaryo ay ibinibigay ng aa. at vv. ovaricae at uterinae. Ang parehong ovarian arteries (aa. ovaricae dextra et sinistra) ay nagmumula sa nauuna na ibabaw ng aorta sa ibaba lamang ng renal arteries; ang kanan ay karaniwang nagmumula sa aorta, at ang kaliwa ay mula sa renal artery. Ang pagdidirekta pababa at sa gilid sa kahabaan ng anterior surface ng psoas major muscle, ang bawat ovarian artery ay tumatawid sa ureter sa harap (nagbibigay ng mga sanga dito), ang mga panlabas na iliac vessel, ang border line at pumapasok sa pelvic cavity, na matatagpuan dito sa suspensory ligament ng ovary. Sumusunod sa medial na direksyon, ang ovarian artery ay dumadaan sa pagitan ng mga dahon ng malawak na ligament ng matris sa ilalim ng fallopian tube, na nagbibigay ng mga sanga dito, at pagkatapos ay sa mesentery ng obaryo; pumapasok ito sa hilum ng obaryo.
Ang mga sanga ng ovarian artery ay malawakang anastomose sa mga ovarian branch ng uterine artery. Ang venous outflow mula sa mga ovary ay isinasagawa pangunahin sa ovarian venous plexus, na matatagpuan sa rehiyon ng ovarian hilum. Mula dito, ang pag-agos ng dugo ay dumadaan sa dalawang direksyon: sa pamamagitan ng uterine at ovarian veins. Ang kanang ovarian vein ay may mga balbula at dumadaloy sa inferior vena cava. Ang kaliwang ovarian vein ay dumadaloy sa kaliwang renal vein, bagaman wala itong mga balbula.
Ang lymphatic drainage mula sa mga ovary ay nangyayari sa pamamagitan ng mga lymphatic vessel, lalo na sa sagana sa lugar ng organ gate, kung saan ang subovarian lymphatic plexus ay nakikilala. Pagkatapos ang lymph ay pinalabas kasama ang mga ovarian lymphatic vessel sa paraaortic lymph nodes.
Innervation ng mga ovary
Sympathetic - ibinibigay ng postganglionic fibers mula sa celiac (solar), superior mesenteric at hypogastric plexuses; parasympathetic - sa pamamagitan ng visceral sacral nerves.
Istraktura ng obaryo
Ang ibabaw ng obaryo ay natatakpan ng isang solong-layer na germinal epithelium. Sa ilalim nito ay namamalagi ang isang siksik na connective tissue protein coat (tunica albuginea). Ang connective tissue ng ovary ay bumubuo ng stroma nito (stroma ovarii), na mayaman sa nababanat na mga hibla. Ang sangkap ng obaryo, ang parenkayma nito, ay nahahati sa panlabas at panloob na mga layer. Ang panloob na layer, na matatagpuan sa gitna ng obaryo, mas malapit sa gate nito, ay tinatawag na medulla (medulla ovarii). Sa layer na ito, sa maluwag na connective tissue, mayroong maraming dugo at lymphatic vessels at nerves. Ang panlabas na layer ng obaryo - ang cortex (cortex ovarii) ay mas siksik. Naglalaman ito ng maraming connective tissue, kung saan matatagpuan ang maturing primary ovarian follicles (folliculi ovarici primarii), pangalawang (vesicular) follicles (folliculi ovarici secundarii, s.vesiculosi), pati na rin ang mature follicles, Graafian follicles (folliculi ovarici maturis), pati na rin ang dilaw at mga katawan.
Ang bawat follicle ay naglalaman ng isang babaeng reproductive ovum, o oocyte (ovocytus). Ang ovum ay hanggang sa 150 μm ang lapad, bilog, naglalaman ng isang nucleus, isang malaking halaga ng cytoplasm, na, bilang karagdagan sa mga cellular organelles, ay naglalaman ng mga protina-lipid inclusions (yolk), glycogen, na kinakailangan para sa nutrisyon ng ovum. Karaniwang nauubos ng ovum ang supply nito ng nutrients sa loob ng 12-24 na oras pagkatapos ng obulasyon. Kung hindi nangyari ang pagpapabunga, ang ovum ay namamatay.
Ang itlog ng tao ay may dalawang lamad na tumatakip dito. Sa loob ay ang cytolemma, na siyang cytoplasmic membrane ng itlog. Sa labas ng cytolemma ay isang layer ng tinatawag na follicular cells, na nagpoprotekta sa itlog at nagsasagawa ng hormone-forming function - naglalabas sila ng estrogens.
Ang physiological na posisyon ng matris, tubes at ovaries ay ibinibigay ng pagsususpinde, pag-aayos at pagsuporta sa mga apparatus na pinag-iisa ang peritoneum, ligaments at pelvic tissue. Ang suspending apparatus ay kinakatawan ng mga nakapares na pormasyon, kabilang dito ang bilog at malawak na ligaments ng matris, tamang ligaments at suspending ligaments ng ovaries. Ang malawak na ligaments ng matris, maayos at suspending ligaments ng mga ovaries ay humahawak sa matris sa gitnang posisyon. Hinihila ng mga bilog na ligament ang fundus ng matris pasulong at nagbibigay ng physiological tilt nito.
Tinitiyak ng apparatus ng pag-aayos (angkla) ang posisyon ng matris sa gitna ng maliit na pelvis at ginagawang halos imposible na lumipat ito sa mga gilid, pabalik-balik. Ngunit dahil ang ligamentous apparatus ay umalis mula sa matris sa mas mababang seksyon nito, posible ang mga tilts ng matris sa iba't ibang direksyon. Kasama sa fixing apparatus ang ligaments na matatagpuan sa maluwag na tissue ng pelvis at umaabot mula sa lower section ng uterus hanggang sa lateral, anterior at posterior wall ng pelvis: sacro-magic, cardinal, uterovesical at vesicopubic ligaments.
Bilang karagdagan sa mesovarium, ang mga sumusunod na ovarian ligaments ay nakikilala:
- ang suspensory ligament ng ovary, na dating itinalaga bilang infundibulopelvic ligament. Ito ay isang fold ng peritoneum na may mga daluyan ng dugo (a. et v. ovarica) at mga lymphatic vessel at nerbiyos ng obaryo na dumadaan dito, na nakaunat sa pagitan ng lateral wall ng pelvis, ang lumbar fascia (sa lugar ng dibisyon ng karaniwang iliac artery sa panlabas at panloob) at ang itaas na (tubal) na dulo ng obaryo;
- ang tamang ligament ng obaryo ay dumadaan sa pagitan ng mga dahon ng malawak na litid ng matris, mas malapit sa posterior leaf, at nag-uugnay sa ibabang dulo ng obaryo sa lateral na gilid ng matris. Ang tamang ligament ng obaryo ay nakakabit sa matris sa pagitan ng simula ng fallopian tube at ng bilog na ligament, sa likod at sa itaas ng huli. Sa kapal ng ligament pass rr. ovarii, na siyang mga terminal na sanga ng uterine artery;
- appendicular-ovarian ligament Ang ligament ay umaabot mula sa mesentery ng apendiks hanggang sa kanang obaryo o malawak na ligament ng matris sa anyo ng isang fold ng peritoneum. Ang ligament ay hindi pare-pareho at sinusunod sa 1/2 - 1/3 ng mga kababaihan.
Ang pagsuporta sa aparato ay kinakatawan ng mga kalamnan at fascia ng pelvic floor, nahahati sa mas mababang, gitna at itaas (panloob) na mga layer.
Ang pinakamalakas ay ang upper (panloob) na layer ng kalamnan, na kinakatawan ng nakapares na kalamnan na nag-aangat sa anus. Binubuo ito ng mga bundle ng kalamnan na lumalabas mula sa coccyx hanggang sa pelvic bone sa tatlong direksyon (pubococcygeus, iliococcygeus, at ischiococcygeus). Ang layer na ito ng mga kalamnan ay tinatawag ding pelvic diaphragm.
Ang gitnang layer ng mga kalamnan ay matatagpuan sa pagitan ng symphysis, pubic at ischial bones. Ang gitnang layer ng mga kalamnan - ang urogenital diaphragm - ay sumasakop sa anterior na kalahati ng pelvic outlet, kung saan dumadaan ang urethra at puki. Sa nauuna na seksyon sa pagitan ng mga sheet nito ay mga bundle ng kalamnan na bumubuo sa panlabas na sphincter ng urethra, sa posterior section ay mga bundle ng kalamnan na papunta sa nakahalang direksyon - ang malalim na nakahalang kalamnan ng perineum.
Ang mas mababang (panlabas) na layer ng pelvic floor muscles ay binubuo ng mga mababaw na kalamnan, ang hugis nito ay kahawig ng numero 8. Kabilang dito ang bulbocavernous, ischiocavernous, external anal sphincter, at superficial transverse perineal muscle.
Ontogenesis ng mga ovary
Ang proseso ng paglaki ng follicle at atresia ay nagsisimula sa 20 linggo ng pagbubuntis, at sa oras ng paghahatid, hanggang sa 2 milyong oocytes ang nananatili sa mga ovary ng batang babae. Sa pamamagitan ng menarche, ang kanilang bilang ay bumababa sa 300 libo. Sa buong panahon ng reproductive life, hindi hihigit sa 500 follicle ang umabot sa maturity at ovulate. Ang paunang paglaki ng mga follicle ay hindi nakasalalay sa pagpapasigla ng FSH, limitado, at mabilis na nangyayari ang atresia. Ito ay pinaniniwalaan na sa halip na mga steroid hormone, ang mga lokal na autocrine/paracrine peptides ang pangunahing regulator ng paglaki at atresia ng mga pangunahing follicle. Ito ay pinaniniwalaan na ang proseso ng paglaki ng follicle at atresia ay hindi naaabala ng anumang mga proseso ng physiological. Ang prosesong ito ay nagpapatuloy sa lahat ng edad, kabilang ang intrauterine period at menopause, at naantala ng pagbubuntis, obulasyon, at anovulation. Ang mekanismo na nagpapalitaw ng paglaki ng follicle at ang kanilang bilang sa bawat tiyak na cycle ay hindi pa rin malinaw.
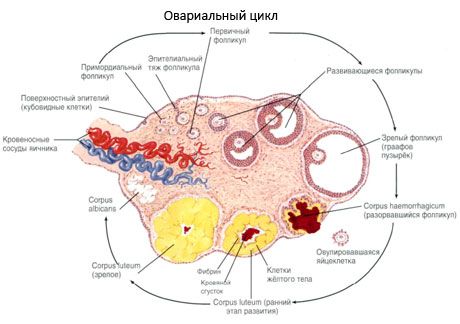
Sa panahon ng pag-unlad nito, ang follicle ay sumasailalim sa ilang mga yugto ng pag-unlad. Ang mga primordial germ cell ay nagmula sa endoderm ng yolk sac, allantois at lumilipat sa genital area ng embryo sa 5-6 na linggo ng pagbubuntis. Bilang resulta ng mabilis na mitotic division, na nagpapatuloy mula 6-8 na linggo hanggang 16-20 na linggo ng pagbubuntis, hanggang sa 6-7 milyong oocytes ang nabuo sa mga ovary ng embryo, na napapalibutan ng manipis na layer ng granulosa cells.
Preantral follicle - ang oocyte ay napapalibutan ng isang lamad (Zona pellucida). Ang mga selulang granulosa na nakapalibot sa oocyte ay nagsisimulang dumami, ang kanilang paglaki ay nakasalalay sa mga gonadotropin at nakakaugnay sa antas ng mga estrogen. Ang mga cell ng Granulosa ay ang target para sa FSH. Sa yugto ng preantral follicle, ang mga granulosa cell ay may kakayahang mag-synthesize ng tatlong klase ng mga steroid: higit sa lahat ay nag-uudyok sa aktibidad ng aromatase, ang pangunahing enzyme na nagko-convert ng androgens sa estradiol. Ito ay pinaniniwalaan na ang estradiol ay may kakayahang dagdagan ang bilang ng sarili nitong mga receptor, na nagbibigay ng direktang mitogenic na epekto sa mga cell ng granulosa na independyente sa FSH. Ito ay itinuturing na paracrine factor na nagpapahusay sa mga epekto ng FSH, kabilang ang pag-activate ng mga proseso ng aromatization.
Lumilitaw ang mga receptor ng FSH sa mga lamad ng mga selula ng granulosa sa sandaling magsimula ang paglaki ng follicle. Ang pagbaba o pagtaas ng FSH ay humahantong sa pagbabago sa bilang ng mga receptor nito. Ang pagkilos na ito ng FSH ay binago ng mga kadahilanan ng paglago. Ang FSH ay kumikilos sa pamamagitan ng G-protein, ang adenylate cyclase system, bagaman ang steroidogenesis sa follicle ay pangunahing kinokontrol ng FSH, maraming mga kadahilanan ang kasangkot sa prosesong ito: mga channel ng ion, tyrosine kinase receptors, ang phospholipase system ng mga pangalawang mensahero.
Ang papel ng androgens sa maagang pag-unlad ng follicle ay kumplikado. Ang mga selulang Granulosa ay may mga androgen receptor. Ang mga ito ay hindi lamang isang substrate para sa FSH-induced aromatization sa mga estrogen, ngunit maaari ring mapahusay ang aromatization sa mababang konsentrasyon. Kapag tumaas ang antas ng androgen, mas pinipili ng mga preantral granulosa cell ang hindi ang aromatization pathway sa estrogens ngunit ang mas simple na androgen pathway sa pamamagitan ng 5a-reductase upang ma-convert sa isang androgen na hindi mako-convert sa estrogen, at sa gayon ay humahadlang sa aktibidad ng aromatase. Pinipigilan din ng prosesong ito ang pagbuo ng FSH at LH receptor, kaya huminto sa pag-unlad ng follicle.
Ang proseso ng aromatization, ang follicle na may mataas na antas ng androgen ay sumasailalim sa mga proseso ng atresia. Ang paglaki at pag-unlad ng follicle ay nakasalalay sa kakayahan nitong i-convert ang androgens sa estrogens.
Sa pagkakaroon ng FSH, ang nangingibabaw na sangkap ng follicular fluid ay mga estrogen. Sa kawalan ng FSH - androgens. Ang LH ay karaniwang wala sa follicular fluid hanggang sa gitna ng cycle. Sa sandaling tumaas ang antas ng LH, bumababa ang mitotic na aktibidad ng mga selula ng granulosa, lumilitaw ang mga degenerative na pagbabago at tumataas ang antas ng androgen sa follicle. Ang antas ng mga steroid sa follicular fluid ay mas mataas kaysa sa plasma at sumasalamin sa functional na aktibidad ng mga ovarian cells: granulosa at theca cells. Kung ang tanging target para sa FSH ay granulosa cells, kung gayon ang LH ay maraming target - theca cells, stromal at lutein cells at granulosa cells. Parehong granulosa at theca cell ay may kakayahan sa steroidogenesis, ngunit ang aromatase activity ay nangingibabaw sa granulosa cells.
Bilang tugon sa LH, ang mga theca cell ay gumagawa ng androgens, na pagkatapos ay na-convert ng mga granulosa cells sa mga estrogen sa pamamagitan ng FSH-induced aromatization.
Habang lumalaki ang follicle, ang mga theca cell ay nagsisimulang magpahayag ng mga gene para sa mga LH receptor, P450 sec at 3beta-hydroxysteroid dehydrogenase, insulin-like growth factor (IGF-1) na synergistically sa LH ay nagpapataas ng gene expression, ngunit hindi nagpapasigla sa steroidogenesis.
Ang ovarian steroidogenesis ay palaging umaasa sa LH. Habang lumalaki ang follicle, ang mga selulang theca ay nagpapahayag ng P450c17 enzyme, na bumubuo ng androgen mula sa kolesterol. Ang mga selulang Granulosa ay walang enzyme na ito at umaasa sa mga selulang theca upang makagawa ng mga estrogen mula sa androgens. Hindi tulad ng steroidogenesis, ang folliculogenesis ay umaasa sa FSH. Habang lumalaki ang follicle at tumataas ang antas ng estrogen, naisaaktibo ang mekanismo ng feedback - pinipigilan ang produksyon ng FSH, na humahantong naman sa pagbaba sa aktibidad ng aromatase ng follicle at, sa huli, sa follicular atresia sa pamamagitan ng apoptosis (programmed cell death).
Ang mekanismo ng feedback ng estrogens at FSH ay pumipigil sa pagbuo ng mga follicle na nagsimulang lumaki, ngunit hindi ang nangingibabaw na follicle. Ang nangingibabaw na follicle ay naglalaman ng higit pang mga FSH receptor, na sumusuporta sa paglaganap ng granulosa cells at ang aromatization ng androgens sa estrogens. Bilang karagdagan, ang paracrine at autocrine pathway ay kumikilos bilang isang mahalagang coordinator ng antral follicle development.
Ang autocrine/paracrine regulator ay binubuo ng mga peptides (inhibin, activin, follistatin), na na-synthesize ng granulosa cells bilang tugon sa FSH at pumapasok sa follicular fluid. Binabawasan ng inhibin ang pagtatago ng FSH; pinasisigla ng activin ang pagpapalabas ng FSH mula sa pituitary gland at pinahuhusay ang pagkilos ng FSH sa obaryo; Pinipigilan ng follistatin ang aktibidad ng FSH, posibleng sa pamamagitan ng nagbubuklod na activin. Pagkatapos ng obulasyon at pag-unlad ng corpus luteum, ang inhibin ay nasa ilalim ng kontrol ng LH.
Ang paglaki at pagkakaiba ng mga ovarian cells ay naiimpluwensyahan ng insulin-like growth factor (IGE). Ang IGF-1 ay kumikilos sa mga cell ng granulosa, na nagdudulot ng pagtaas sa cyclic adenosine monophosphate (cAMP), progesterone, oxytocin, proteoglycan, at inhibin.
Ang IGF-1 ay kumikilos sa theca cells, na nagiging sanhi ng pagtaas ng produksyon ng androgen. Ang mga selula ng theca, sa turn, ay gumagawa ng tumor necrosis factor (TNF) at epidermal growth factor (EGF), na kinokontrol din ng FSH.
Pinasisigla ng EGF ang paglaganap ng mga selulang granulosa. Ang IGF-2 ay ang pangunahing kadahilanan ng paglago sa follicular fluid, at ang IGF-1, TNF-a, TNF-3, at EGF ay matatagpuan din dito.
Ang pagkagambala sa paracrine at/o autocrine regulation ng ovarian function ay lumilitaw na gumaganap ng isang papel sa obulasyon disorder at sa pagbuo ng polycystic ovaries.
Habang lumalaki ang antral follicle, tumataas ang estrogen content sa follicular fluid. Sa tuktok ng kanilang pagtaas, lumilitaw ang mga receptor para sa LH sa mga selula ng granulosa, nangyayari ang luteinization ng mga selulang granulosa, at tumataas ang produksyon ng progesterone. Kaya, sa preovultory period, ang pagtaas sa produksyon ng estrogen ay nagiging sanhi ng paglitaw ng mga LH receptors, ang LH naman, ay nagiging sanhi ng luteinization ng granulosa cells at progesterone production. Ang pagtaas sa progesterone ay binabawasan ang mga antas ng estrogen, na tila nagiging sanhi ng pangalawang peak ng FSH sa gitna ng cycle.
Ang obulasyon ay naisip na mangyari 10-12 oras pagkatapos ng LH peak at 24-36 na oras pagkatapos ng estradiol peak. Ang LH ay naisip na pasiglahin ang pagbawas ng dibisyon ng oocyte, luteinization ng granulosa cells, at synthesis ng progesterone at prostaglandin sa follicle.
Pinahuhusay ng progesterone ang aktibidad ng mga proteolytic enzymes, na kasama ng prostaglandin ay lumahok sa pagkalagot ng follicle wall. Ang progesterone-induced peak ng FSH ay nagpapahintulot sa oocyte na lumabas sa follicle sa pamamagitan ng pag-convert ng plasminogen sa proteolytic enzyme plasmin, at nagbibigay ng sapat na bilang ng LH receptors para sa normal na pag-unlad ng luteal phase.
Sa loob ng 3 araw pagkatapos ng obulasyon, tumataas ang laki ng mga selula ng granulosa, at ang mga katangian ng vacuole na puno ng pigment, lutein, ay lilitaw sa kanila. Ang mga selulang theca-luteal ay naiiba sa theca at stroma at naging bahagi ng corpus luteum. Ang mga capillary na tumagos sa corpus luteum ay mabilis na umuunlad sa ilalim ng impluwensya ng mga kadahilanan ng angiogenesis, at sa pinabuting vascularization, ang produksyon ng progesterone at estrogen ay tumataas. Ang aktibidad ng steroidogenesis at ang habang-buhay ng corpus luteum ay tinutukoy ng antas ng LH. Ang corpus luteum ay hindi isang homogenous na cellular formation. Bilang karagdagan sa 2 uri ng luteal cells, naglalaman ito ng mga endothelial cells, macrophage, fibroblast, atbp. Ang malalaking luteal cells ay gumagawa ng mga peptides (relaxin, oxytocin) at mas aktibo sa steroidogenesis na may mas malaking aktibidad ng aromatase at mas malaking synthesis ng progesterone kaysa sa maliliit na selula.
Ang peak ng progesterone ay nangyayari sa ika-8 araw pagkatapos ng LH peak. Nabanggit na ang progesterone at estradiol ay itinago nang episodically sa luteal phase na may kaugnayan sa pulsatile na output ng LH. Sa pagbuo ng corpus luteum, ang kontrol ng produksyon ng inhibin ay pumasa mula sa FSH hanggang LH. Ang inhibin ay tumataas sa pagtaas ng estradiol bago ang LH peak at patuloy na tumataas pagkatapos ng LH peak, kahit na ang mga antas ng estrogen ay bumababa. Bagaman ang inhibin at estradiol ay itinago ng mga selula ng granulosa, kinokontrol sila ng iba't ibang mga landas. Ang pagbaba ng inhibin sa pagtatapos ng luteal phase ay nag-aambag sa pagtaas ng FSH para sa susunod na cycle.
Ang corpus luteum ay bumaba nang napakabilis - sa ika-9-11 araw pagkatapos ng obulasyon.
Ang mekanismo ng pagkabulok ay hindi malinaw at hindi nauugnay sa luteolytic na papel ng mga estrogen o sa isang mekanismo na nauugnay sa receptor, tulad ng naobserbahan sa endometrium. May isa pang paliwanag para sa papel ng mga estrogen na ginawa ng corpus luteum. Ito ay kilala na ang mga estrogen ay kinakailangan para sa synthesis ng progesterone receptors sa endometrium. Ang mga luteal phase estrogen ay malamang na kinakailangan para sa mga pagbabagong nauugnay sa progesterone sa endometrium pagkatapos ng obulasyon. Ang hindi sapat na pag-unlad ng mga receptor ng progesterone, bilang resulta ng hindi sapat na antas ng estrogen, ay posibleng isang karagdagang mekanismo para sa kawalan ng katabaan at pagkawala ng maagang pagbubuntis, isa pang anyo ng kakulangan sa luteal phase. Ito ay pinaniniwalaan na ang haba ng buhay ng corpus luteum ay itinatag sa oras ng obulasyon. At tiyak na babalik ito kung hindi ito susuportahan ng human chorionic gonadotropin dahil sa pagbubuntis. Kaya, ang regression ng corpus luteum ay humahantong sa pagbaba sa mga antas ng estradiol, progesterone at inhibin. Ang pagbaba sa inhibin ay nag-aalis ng suppressive effect nito sa FSH; ang pagbaba sa estradiol at progesterone ay nagbibigay-daan para sa isang napakabilis na pagpapanumbalik ng pagtatago ng GnRH at ang pag-alis ng mekanismo ng feedback mula sa pituitary gland. Ang pagbaba ng inhibin at estradiol, kasama ang pagtaas ng GnRH, ay nagbubunga ng FSH sa LH. Ang pagtaas ng FSH ay humahantong sa paglaki ng follicle na may kasunod na pagpili ng isang nangingibabaw na follicle, at magsisimula ang isang bagong cycle kung hindi nangyari ang pagbubuntis. Ang mga steroid na hormone ay gumaganap ng isang nangungunang papel sa reproductive biology at pangkalahatang pisyolohiya. Tinutukoy nila ang phenotype ng tao, nakakaapekto sa cardiovascular system, metabolismo ng buto, balat, pangkalahatang kagalingan at may mahalagang papel sa pagbubuntis. Ang pagkilos ng mga steroid hormones ay sumasalamin sa intracellular at genetic na mga mekanismo na kinakailangan upang magpadala ng extracellular signal sa cell nucleus upang magdulot ng physiological response.
Ang mga estrogen ay diffusely tumagos sa cell lamad at nagbubuklod sa mga receptor na matatagpuan sa cell nucleus. Ang receptor-steroid complex pagkatapos ay nagbubuklod sa DNA. Sa mga target na cell, ang mga pakikipag-ugnayan na ito ay humahantong sa pagpapahayag ng gene, synthesis ng protina, at tiyak na paggana ng cell at tissue.