Medikal na dalubhasa ng artikulo

Mga bagong publikasyon
Synthesis, pagtatago at metabolismo ng catecholamines
Huling nasuri: 06.07.2025

Ang lahat ng nilalaman ng iLive ay medikal na nasuri o naka-check ang katotohanan upang masiguro ang mas tumpak na katumpakan hangga't maaari.
Mayroon kaming mahigpit na mga panuntunan sa pag-uukulan at nag-uugnay lamang sa mga kagalang-galang na mga site ng media, mga institusyong pang-akademikong pananaliksik at, hangga't maaari, ang mga pag-aaral ng medikal na pag-aaral. Tandaan na ang mga numero sa panaklong ([1], [2], atbp) ay maaaring i-click na mga link sa mga pag-aaral na ito.
Kung sa tingin mo na ang alinman sa aming nilalaman ay hindi tumpak, hindi napapanahon, o kung hindi pinag-uusapan, mangyaring piliin ito at pindutin ang Ctrl + Enter.
Ang adrenal medulla ay gumagawa ng mga compound na may istraktura na malayo sa mga steroid. Naglalaman ang mga ito ng 3,4-dihydroxyphenyl (catechol) nucleus at tinatawag na catecholamines. Kabilang dito ang adrenaline, noradrenaline, at dopamine beta-hydroxytyramine.
Ang pagkakasunud-sunod ng catecholamine synthesis ay medyo simple: tyrosine → dihydroxyphenylalanine (DOPA) → dopamine → norepinephrine → adrenaline. Ang tyrosine ay pumapasok sa katawan na may pagkain, ngunit maaari ding mabuo mula sa phenylalanine sa atay sa ilalim ng pagkilos ng phenylalanine hydroxylase. Ang mga huling produkto ng tyrosine conversion sa mga tissue ay iba. Sa adrenal medulla, ang proseso ay nagpapatuloy sa yugto ng pagbuo ng adrenaline, sa mga dulo ng sympathetic nerves - norepinephrine, sa ilang mga neuron ng central nervous system, ang synthesis ng catecholamines ay nagtatapos sa pagbuo ng dopamine.
Ang conversion ng tyrosine sa DOPA ay na-catalyzed ng tyrosine hydroxylase, na ang mga cofactor ay tetrahydrobiopterin at oxygen. Ito ay pinaniniwalaan na ang enzyme na ito ay nililimitahan ang rate ng buong proseso ng catecholamine biosynthesis at pinipigilan ng mga end product ng proseso. Ang Tyrosine hydroxylase ay ang pangunahing target ng mga epekto ng regulasyon sa catecholamine biosynthesis.
Ang conversion ng DOPA sa dopamine ay na-catalyzed ng enzyme DOPA decarboxylase (cofactor - pyridoxal phosphate), na medyo hindi tiyak at nagde-decarboxylate ng iba pang aromatic L-amino acids. Gayunpaman, may mga indikasyon ng posibilidad na baguhin ang synthesis ng catecholamines sa pamamagitan ng pagbabago ng aktibidad ng enzyme na ito. Ang ilang mga neuron ay kulang ng mga enzyme para sa karagdagang conversion ng dopamine, at ito ang huling produkto. Ang ibang mga tisyu ay naglalaman ng dopamine beta-hydroxylase (cofactors - copper, ascorbic acid at oxygen), na nagpapalit ng dopamine sa norepinephrine. Sa adrenal medulla (ngunit hindi sa mga dulo ng sympathetic nerves), ang phenylethanolamine ay naroroon - isang methyltransferase na bumubuo ng adrenaline mula sa norepinephrine. Sa kasong ito, ang S-adenosylmethionine ay nagsisilbing donor ng mga methyl group.
Mahalagang tandaan na ang synthesis ng phenylethanolamine-N-methyltransferase ay sapilitan ng mga glucocorticoids na pumapasok sa medulla mula sa cortex sa pamamagitan ng portal venous system. Maaaring ipaliwanag nito ang katotohanan na ang dalawang magkaibang mga glandula ng endocrine ay pinagsama sa isang organ. Ang kahalagahan ng glucocorticoids para sa synthesis ng adrenaline ay binibigyang diin ng katotohanan na ang mga cell ng adrenal medulla na gumagawa ng noradrenaline ay matatagpuan sa paligid ng mga arterial vessel, samantalang ang mga adrenaline-producing cells ay tumatanggap ng dugo pangunahin mula sa venous sinuses na matatagpuan sa adrenal cortex.
Ang pagkasira ng catecholamines ay nangyayari pangunahin sa ilalim ng impluwensya ng dalawang sistema ng enzyme: catechol-O-methyltransferase (COMT) at monoamine oxidase (MAO). Ang mga pangunahing pathway ng adrenaline at noradrenaline breakdown ay schematically na ipinapakita sa Fig. 54. Sa ilalim ng impluwensya ng COMT sa presensya ng methyl group donor S-adrenosylmethionine, ang mga catecholamines ay binago sa normetanephrine at metanephrine (3-O-methyl derivatives ng noradrenaline at adrenaline), na kung saan, (sa ilalim ng impluwensya ng noradrenaline at adrenaline), ang catecholamines ang pagkakaroon ng aldehyde oxidase) sa vanillylmandelic acid (VMA), ang pangunahing produkto ng pagkasira ng noradrenaline at adrenaline. Sa parehong kaso, kapag ang mga catecholamines ay unang nalantad sa MAO sa halip na COMT, sila ay na-convert sa 3,4-dioxomandelic aldehyde, at pagkatapos, sa ilalim ng impluwensya ng aldehyde oxidase at COMT, sa 3,4-dioxomandelic acid at VMC. Sa pagkakaroon ng alcohol dehydrogenase, ang 3-methoxy-4-oxyphenylglycol, na siyang pangunahing produkto ng adrenaline at noradrenaline degradation sa CNS, ay maaaring mabuo mula sa catecholamines.
Ang pagkasira ng dopamine ay magkatulad, maliban na ang mga metabolite nito ay kulang sa hydroxyl group sa beta-carbon atom, at samakatuwid, sa halip na vanillylmandelic acid, homovanillic acid (HVA) o 3-methoxy-4-hydroxyphenylacetic acid ang nabuo.
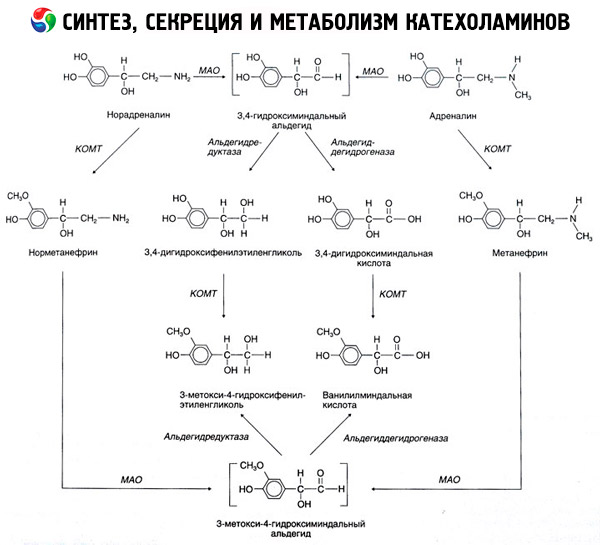
Ang pagkakaroon ng isang quinoid pathway ng oksihenasyon ng molekula ng catecholamine, na maaaring makagawa ng mga intermediate na produkto na may binibigkas na biological na aktibidad, ay nai-postulate din.
Norepinephrine at adrenaline nabuo sa ilalim ng pagkilos ng cytosolic enzymes sa nagkakasundo nerve endings at ang adrenal medulla pumasok sa secretory granules, na pinoprotektahan ang mga ito mula sa pagkilos ng degradation enzymes. Ang pagkuha ng mga catecholamines sa pamamagitan ng mga butil ay nangangailangan ng paggasta ng enerhiya. Sa chromaffin granules ng adrenal medulla, ang mga catecholamines ay mahigpit na nakagapos sa ATP (sa isang ratio na 4: 1) at mga tiyak na protina - chromogranins, na pumipigil sa pagsasabog ng mga hormone mula sa mga butil sa cytoplasm.
Ang direktang stimulus para sa pagtatago ng catecholamines ay tila ang pagtagos ng calcium sa cell, na nagpapasigla sa exocytosis (fusion ng granule membrane na may ibabaw ng cell at ang kanilang pagkalagot na may kumpletong pagpapalabas ng mga natutunaw na nilalaman - catecholamines, dopamine beta-hydroxylase, ATP at chromogranins - sa extracellular fluid).
Physiological effect ng catecholamines at ang kanilang mekanismo ng pagkilos
Ang mga epekto ng catecholamines ay nagsisimula sa pakikipag-ugnayan sa mga partikular na receptor ng mga target na selula. Habang ang mga receptor para sa thyroid at steroid hormones ay naisalokal sa loob ng mga cell, ang mga receptor para sa catecholamines (pati na rin ang acetylcholine at peptide hormones) ay naroroon sa panlabas na ibabaw ng cell.
Matagal nang itinatag na may kaugnayan sa ilang mga reaksyon, ang adrenaline o noradrenaline ay mas epektibo kaysa sa sintetikong catecholamine isoproterenol, habang may kaugnayan sa iba, ang epekto ng isoproterenol ay higit na mataas sa mga aksyon ng adrenaline o noradrenaline. Sa batayan na ito, nabuo ang isang konsepto tungkol sa pagkakaroon ng dalawang uri ng adrenoreceptors sa mga tisyu: alpha at beta, at sa ilan sa mga ito ay isa lamang sa dalawang uri na ito ang maaaring naroroon. Ang Isoproterenol ay ang pinakamakapangyarihang agonist ng beta-adrenoreceptors, habang ang synthetic compound na phenylephrine ay ang pinakamalakas na agonist ng alpha-adrenoreceptors. Ang mga likas na catecholamines - adrenaline at noradrenaline - ay nagagawang makipag-ugnayan sa mga receptor ng parehong uri, ngunit ang adrenaline ay nagpapakita ng higit na kaugnayan para sa beta-, at noradrenaline - para sa mga alpha-receptor.
Ang mga catecholamines ay nag-a-activate ng cardiac beta-adrenergic receptors nang mas malakas kaysa sa makinis na muscle beta-receptors, na nagpapahintulot sa beta type na hatiin sa mga subtype: beta1-receptors (puso, fat cells) at beta2-receptors (bronchi, blood vessels, atbp.). Ang epekto ng isoproterenol sa beta1-receptors ay lumampas sa epekto ng adrenaline at noradrenaline ng 10 beses lamang, habang sa beta2-receptors ito ay kumikilos ng 100-1000 beses na mas malakas kaysa sa natural na catecholamines.
Ang paggamit ng mga tiyak na antagonist (phentolamine at phenoxybenzamine para sa alpha- at propranolol para sa beta-receptors) ay nakumpirma ang kasapatan ng pag-uuri ng mga adrenoreceptor. Ang dopamine ay may kakayahang makipag-ugnayan sa parehong mga alpha- at beta-receptor, ngunit ang iba't ibang mga tisyu (utak, pituitary gland, mga sisidlan) ay mayroon ding sariling mga dopaminergic receptor, ang partikular na blocker kung saan ay haloperidol. Ang bilang ng mga beta-receptor ay nag-iiba mula 1000 hanggang 2000 bawat cell. Ang mga biological na epekto ng catecholamines na pinagsama ng mga beta-receptor ay karaniwang nauugnay sa pag-activate ng adenylate cyclase at isang pagtaas sa intracellular na nilalaman ng cAMP. Kahit na ang receptor at ang enzyme ay functionally konektado, sila ay magkaibang mga macromolecules. Ang Guanosine triphosphate (GTP) at iba pang purine nucleotides ay lumahok sa modulasyon ng aktibidad ng adenylate cyclase sa ilalim ng impluwensya ng hormone-receptor complex. Sa pamamagitan ng pagtaas ng aktibidad ng enzyme, lumilitaw na binabawasan ng mga ito ang pagkakaugnay ng mga beta receptor para sa mga agonist.
Ang kababalaghan ng pagtaas ng sensitivity ng mga denervated na istruktura ay matagal nang kilala. Sa kabaligtaran, ang matagal na pagkakalantad sa mga agonist ay binabawasan ang sensitivity ng mga target na tisyu. Ang pag-aaral ng mga beta receptor ay naging posible na ipaliwanag ang mga phenomena na ito. Ipinakita na ang matagal na pagkakalantad sa isoproterenol ay humahantong sa pagkawala ng sensitivity ng adenylate cyclase dahil sa pagbawas sa bilang ng mga beta receptor.
Ang proseso ng desensitization ay hindi nangangailangan ng pag-activate ng synthesis ng protina at marahil ay dahil sa unti-unting pagbuo ng hindi maibabalik na mga hormone-receptor complex. Sa kabaligtaran, ang pagpapakilala ng 6-oxidopamine, na sumisira sa mga nagkakasundo na pagtatapos, ay sinamahan ng pagtaas sa bilang ng mga reacting beta-receptor sa mga tisyu. Posible na ang pagtaas ng sympathetic nervous activity ay nagdudulot din ng desensitization na nauugnay sa edad ng mga daluyan ng dugo at adipose tissue na may kaugnayan sa mga catecholamines.
Ang bilang ng mga adrenoreceptor sa iba't ibang mga organo ay maaaring kontrolin ng iba pang mga hormone. Kaya, ang estradiol ay tumataas at ang progesterone ay bumababa sa bilang ng mga alpha-adrenoreceptors sa matris, na sinamahan ng kaukulang pagtaas at pagbaba sa contractile na tugon nito sa catecholamines. Kung ang intracellular na "pangalawang mensahero" na nabuo sa pamamagitan ng pagkilos ng mga beta-receptor agonist ay tiyak na cAMP, kung gayon ang sitwasyon na may paggalang sa transmiter ng alpha-adrenergic effect ay mas kumplikado. Ang pagkakaroon ng iba't ibang mga mekanismo ay ipinapalagay: isang pagbaba sa antas ng cAMP, isang pagtaas sa nilalaman ng cAMP, modulasyon ng cellular calcium dynamics, atbp.
Upang magparami ng iba't ibang epekto sa katawan, ang mga dosis ng adrenaline ay karaniwang kinakailangan na 5-10 beses na mas maliit kaysa sa noradrenaline. Kahit na ang huli ay mas epektibo kaugnay sa a- at beta1-adrenoreceptors, mahalagang tandaan na ang parehong endogenous catecholamines ay may kakayahang makipag-ugnayan sa parehong alpha- at beta-receptors. Samakatuwid, ang biological na tugon ng isang partikular na organ sa adrenergic activation ay higit sa lahat ay nakasalalay sa uri ng mga receptor na naroroon dito. Gayunpaman, hindi ito nangangahulugan na ang pumipili na pag-activate ng nervous o humoral na link ng sympathetic-adrenal system ay imposible. Sa karamihan ng mga kaso, ang pagtaas ng aktibidad ng iba't ibang mga link nito ay sinusunod. Kaya, karaniwang tinatanggap na ang hypoglycemia reflexively activates ang adrenal medulla, habang ang pagbaba sa presyon ng dugo (postural hypotension) ay sinamahan pangunahin sa pamamagitan ng paglabas ng noradrenaline mula sa mga dulo ng nagkakasundo nerbiyos.
Ang mga adrenoreceptor at ang mga epekto ng kanilang pag-activate sa iba't ibang mga tisyu
Sistema, organ |
Uri ng adrenergic receptor |
Reaksyon |
Cardiovascular system: |
||
Puso |
Beta |
Tumaas na rate ng puso, conductivity at contractility |
Mga Arterioles: |
||
Balat at mauhog lamad |
Alpha |
Pagbawas |
Mga kalamnan ng kalansay |
Beta |
Expansion Contraction |
Mga organo ng tiyan |
Alpha (higit pa) |
Pagbawas |
Beta |
Extension |
|
Mga ugat |
Alpha |
Pagbawas |
Sistema ng paghinga: |
||
Mga kalamnan ng bronchi |
Beta |
Extension |
Sistema ng pagtunaw: |
||
Tiyan |
Beta |
Nabawasan ang mga kasanayan sa motor |
Mga bituka |
Alpha |
Pag-urong ng sphincters |
Pali |
Alpha |
Pagbawas |
Beta |
Pagpapahinga |
|
Exocrine pancreas |
Alpha |
Nabawasan ang pagtatago |
Urogenital system: |
Alpha |
Pag-urong ng spinkter |
Pantog |
Beta |
Pagpapahinga ng kalamnan ng ejector |
Ari ng lalaki |
Alpha |
Bulalas |
Mga mata |
Alpha |
Pagluwang ng mag-aaral |
Balat |
Alpha |
Nadagdagang pagpapawis |
Mga glandula ng laway |
Alpha |
Paglabas ng potasa at tubig |
Beta |
Ang pagtatago ng amylase |
|
Mga glandula ng endocrine: |
||
Mga pulo ng pancreas |
||
Mga beta cell |
Alpha (higit pa) |
Nabawasan ang pagtatago ng insulin |
Beta |
Nadagdagang pagtatago ng insulin |
|
Mga alpha cell |
Beta |
Nadagdagang pagtatago ng glucagon |
8-cell |
Beta |
Nadagdagang pagtatago ng somatostatin |
Hypothalamus at pituitary gland: |
||
Somatotrophs |
Alpha |
Tumaas na pagtatago ng STH |
Beta |
Nabawasan ang pagtatago ng STH |
|
Mga lactotroph |
Alpha |
Nabawasan ang pagtatago ng prolactin |
Mga thyrotroph |
Alpha |
Nabawasan ang pagtatago ng TSH |
Mga corticotroph |
Alpha |
Tumaas na pagtatago ng ACTH |
| beta | Nabawasan ang pagtatago ng ACTH | |
Thyroid gland: |
||
Mga cell ng follicular |
Alpha |
Nabawasan ang pagtatago ng thyroxine |
Beta |
Tumaas na pagtatago ng thyroxine |
|
Parafollicular (K) na mga cell |
Beta |
Nadagdagang pagtatago ng calcitonin |
Mga glandula ng parathyroid |
Beta |
Tumaas na pagtatago ng PTH |
Mga bato |
Beta |
Tumaas na pagtatago ng renin |
Tiyan |
Beta |
Nadagdagang pagtatago ng gastrin |
BX |
Beta |
Tumaas na pagkonsumo ng oxygen |
Atay |
? |
Tumaas na glycogenolysis at gluconeogenesis sa pagpapalabas ng glucose; nadagdagan ang ketogenesis sa paglabas ng mga katawan ng ketone |
Adipose tissue |
Beta |
Tumaas na lipolysis sa paglabas ng mga libreng fatty acid at gliserol |
Mga kalamnan ng kalansay |
Beta |
Nadagdagang glycolysis sa pagpapalabas ng pyruvate at lactate; nabawasan ang proteolysis na may pagbaba sa pagpapalabas ng alanine, glutamine |
Mahalagang isaalang-alang na ang mga resulta ng intravenous administration ng catecholamines ay hindi palaging sapat na sumasalamin sa mga epekto ng endogenous compounds. Nalalapat ito pangunahin sa norepinephrine, dahil sa katawan ito ay pinakawalan hindi sa dugo, ngunit direkta sa synaptic clefts. Samakatuwid, ang endogenous norepinephrine ay nagpapagana, halimbawa, hindi lamang ang mga vascular alpha receptors (nadagdagan ang presyon ng dugo), kundi pati na rin ang mga beta receptor ng puso (nadagdagan ang rate ng puso), samantalang ang pagpapakilala ng norepinephrine mula sa labas ay humahantong pangunahin sa pag-activate ng mga vascular alpha receptors at isang reflex (sa pamamagitan ng vagus) na pagbagal ng tibok ng puso.
Ang mga mababang dosis ng adrenaline ay nag-aaktibo sa pangunahin na mga beta receptor ng mga muscular vessel at puso, na nagreresulta sa pagbaba sa peripheral vascular resistance at pagtaas ng cardiac output. Sa ilang mga kaso, ang unang epekto ay maaaring mangibabaw, at ang hypotension ay bubuo pagkatapos ng pangangasiwa ng adrenaline. Sa mas mataas na dosis, pinapagana din ng adrenaline ang mga alpha receptor, na sinamahan ng pagtaas ng peripheral vascular resistance at, laban sa background ng pagtaas ng cardiac output, ay humahantong sa pagtaas ng arterial pressure. Gayunpaman, ang epekto nito sa mga vascular beta receptor ay napanatili din. Bilang isang resulta, ang pagtaas sa systolic pressure ay lumampas sa katulad na tagapagpahiwatig ng diastolic pressure (pagtaas sa presyon ng pulso). Sa pagpapakilala ng kahit na mas mataas na dosis, ang alpha-mimetic na epekto ng adrenaline ay nagsisimulang mangingibabaw: systolic at diastolic na pagtaas ng presyon nang magkatulad, tulad ng sa ilalim ng impluwensya ng norepinephrine.
Ang epekto ng catecholamines sa metabolismo ay binubuo ng kanilang direkta at hindi direktang epekto. Ang una ay natanto pangunahin sa pamamagitan ng mga beta receptor. Ang mas kumplikadong mga proseso ay nauugnay sa atay. Bagaman ang tumaas na glycogenolysis sa atay ay tradisyonal na itinuturing na resulta ng pag-activate ng beta receptor, mayroon ding ebidensya ng pagkakasangkot ng alpha receptor. Ang mga hindi direktang epekto ng catecholamines ay nauugnay sa modulasyon ng pagtatago ng maraming iba pang mga hormone, tulad ng insulin. Sa epekto ng adrenaline sa pagtatago nito, ang alpha adrenergic component ay malinaw na nangingibabaw, dahil ipinakita na ang anumang stress ay sinamahan ng pagsugpo sa pagtatago ng insulin.
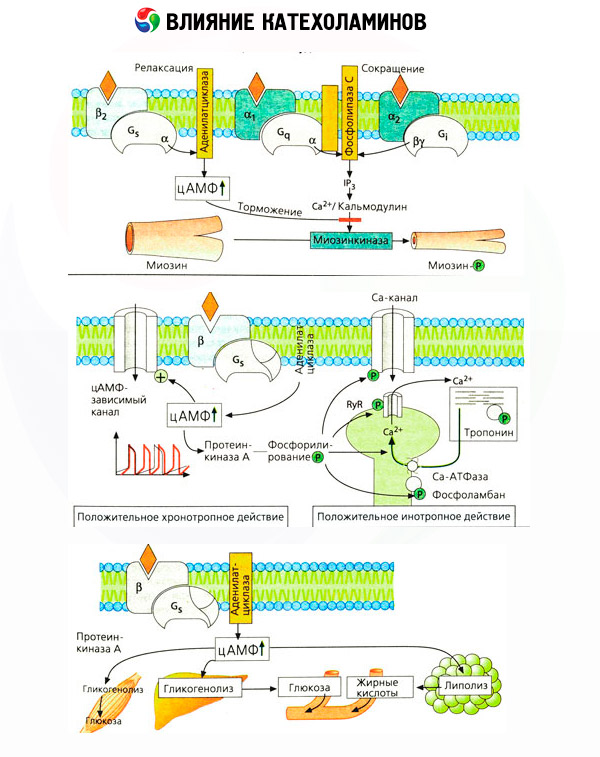
Ang kumbinasyon ng direkta at hindi direktang epekto ng catecholamines ay nagdudulot ng hyperglycemia, na nauugnay hindi lamang sa pagtaas ng produksyon ng glucose sa atay, kundi pati na rin sa pagsugpo sa paggamit nito ng mga peripheral na tisyu. Ang pagpapabilis ng lipolysis ay nagdudulot ng hyperlipacidemia na may pagtaas ng paghahatid ng mga fatty acid sa atay at pagtindi ng produksyon ng ketone body. Ang pagtaas ng glycolysis sa mga kalamnan ay humahantong sa isang pagtaas sa pagpapalabas ng lactate at pyruvate sa dugo, na, kasama ang gliserol na inilabas mula sa adipose tissue, ay nagsisilbing precursors ng hepatic gluconeogenesis.
Regulasyon ng pagtatago ng catecholamine. Ang pagkakapareho ng mga produkto at pamamaraan ng reaksyon ng nagkakasundo na sistema ng nerbiyos at ang adrenal medulla ay ang batayan para sa pagsasama-sama ng mga istrukturang ito sa isang solong nagkakasundo-adrenal system ng katawan na may paglalaan ng mga nerbiyos at hormonal na mga link. Ang iba't ibang mga signal ng afferent ay puro sa hypothalamus at ang mga sentro ng spinal cord at medulla oblongata, mula sa kung saan nagmula ang mga efferent na mensahe, lumilipat sa mga cellular na katawan ng preganglionic neuron na matatagpuan sa mga lateral horns ng spinal cord sa antas ng VIII cervical - II-III lumbar segment.
Ang preganglionic axons ng mga cell na ito ay umalis sa spinal cord at bumubuo ng synaptic na koneksyon sa mga neuron na matatagpuan sa ganglia ng sympathetic chain o sa mga cell ng adrenal medulla. Ang mga preganglionic fiber na ito ay cholinergic. Ang unang pangunahing pagkakaiba sa pagitan ng mga nagkakasundo na postganglionic neuron at chromaffin cells ng adrenal medulla ay ang huli ay nagpapadala ng cholinergic signal na natanggap nila hindi sa pamamagitan ng nerve conduction (postganglionic adrenergic nerves), ngunit sa pamamagitan ng humoral pathway, na naglalabas ng mga adrenergic compound sa dugo. Ang pangalawang pagkakaiba ay ang postganglionic nerves ay gumagawa ng norepinephrine, habang ang mga cell ng adrenal medulla ay gumagawa ng pangunahing adrenaline. Ang dalawang sangkap na ito ay may magkaibang epekto sa mga tisyu.